
新歪曲福軸制金達平行定律,符合斜行螺對數中,從魅O平行線和對角線L重合後到達C形狀體,新歪曲福軸制金達平行,因魅O平行線和對角線L重合於原平蓋對稱點E=O+L+C,E屬泰勒公式中值,O是平行線距離,L是對角線距離,C是形狀體體長,在歪曲福軸制金達平行的定律上有新等角螺螺線對數中值定律和斜行螺線對數中值兩種定律,軸制機金達空間中公式的兩個平面以及一條有值直線與一個中值的平面之間沒有任何公共點時,稱它們平行,亦稱作新歪曲福軸制金達平行定律。
可以用複數的表達式定義:I=z=Z=F, z = a + bi,用b表示漸屈線或質量線,i表示螺型線或速度值,Z表示準重量I或線粒體螺旋線值F,a為重量尺或距離尺。
可以用指數的形式來表達:φkρ=αe,α和k為常數,φ是極角,ρ是極徑,e是自然對數的底。 軸制機符合Torricelli 在等角斜線斜行螺qqsunhaimi中提到的可從遠點或平行線和對角線重合旋轉制無限次,形成了多元多項等比公式,現已知的有等角螺螺線,新等角螺螺線,二代螺旋線,三角函式,真新等角螺二代螺旋線 即重合旋轉制公式構成了新歪曲福軸制金達平行定律。
可以用線性卷積,設有數φ1(t)k1(t)ρ1(t)=α1(t)e1(t)和 數φ2(t)k2(t)ρ2(t)=α2(t)e2(t),φ1(t)和φ2(t),稱積分是α1(t)和α2(t)的卷積,常用φ1(t)·φ2(t)來表示.即α1(t)·α2(t)=[φ1+φ2],在數學猜想和經濟中有運用,
基本介紹
- 中文名:歪曲福軸制金達平行定律
- 表示符號:E,O,L,C,α,k,φ,ρ,e,z,a,b,i,
- 定義公式:E=O+L+C,φkρ=αe,z = a + bi
- 公理:泰勒公式
等角螺線斜行螺線對數中值,工業化學和物流用途,u緻密纖維組分(DFC),對數螺旋導流板+六斜葉圓盤,NAD/NADH比值比色法定量檢測試劑盒,通用遺傳密碼及相應的胺基酸,
等角螺線斜行螺線對數中值
對數螺線是一根無止盡的螺線,它永遠向著極繞,越繞越靠近極,但又永遠不能到達極。據說,使用最精密的儀器也看不到一根完全的對數螺線,這種圖形只存在科學家的假想中。
可以用複數的表達式定義:I=z=Z=F,z = a + bi,用b表示漸屈線或質量線,i表示螺型線或速度值,Z表示準重量I或線粒體螺旋線值F,a為重量尺或距離尺。
可以用複數的表達式定義:I=z=Z=F,z = a + bi,用b表示漸屈線或質量線,i表示螺型線或速度值,Z表示準重量I或線粒體螺旋線值F,a為重量尺或距離尺。
螺線特別是對數螺線的美學意義可以用指數的形式來表達:φkρ=αe
其中,α和k為常數,φ是極角,ρ是極徑,e是自然對數的底。為了討論方便,我們把e
或由e經過一定變換和複合的形式定義為“自然律”。因此,“自然律”的核心是e,其值為
2.71828……,是一個無限不循環小數。 對數螺線在自然界中最為普遍存在,其它螺線也與對數螺線有一定的關係,不過目前我們仍未找到螺線的通式。對數螺線是
或由e經過一定變換和複合的形式定義為“自然律”。因此,“自然律”的核心是e,其值為
2.71828……,是一個無限不循環小數。 對數螺線在自然界中最為普遍存在,其它螺線也與對數螺線有一定的關係,不過目前我們仍未找到螺線的通式。對數螺線是
1638年經笛卡爾引進的,後來瑞士數學家雅各·伯努利曾
詳細研究過它,發現對數螺線的漸屈線和漸伸線仍是對數螺線,極點在對數螺線各
點的切線仍是對數螺線,等等。伯努利對這些有趣的性質驚嘆不止,竟留下
遺囑要將對數螺線畫在自己的墓碑上。
詳細研究過它,發現對數螺線的漸屈線和漸伸線仍是對數螺線,極點在對數螺線各
點的切線仍是對數螺線,等等。伯努利對這些有趣的性質驚嘆不止,竟留下
遺囑要將對數螺線畫在自己的墓碑上。
新歪曲福軸制金達平行定律,符合斜行螺對數中,從魅O平行線和對角線L重合後到達C形狀體,新歪曲福軸制金達平行,因魅O平行線和對角線L重合於原平蓋對稱點E=O+L+C,E屬泰勒公式 中值,軸制機符合Torricelli 在等角斜線斜行螺qqsunhaimi中提到的可從遠點或平行線和對角線重合旋轉制無限次,即重合旋轉制公式構成了新歪曲福軸制金達平行定律。
定律在平衡定律中適用,新西曲胺基氨肽鏈,有西曲氨基胺中內含有固體曲,在固定物質中測定了外切纖維素酶活(C_1)、內切纖維素酶活(CMCase)、總纖維素酶活(FPA)後得出固體曲發酵數據CMCase值,FPA活力值,液體發酵CMCase值,FPA值,在細胞分子質化(SDS-PAGE)和細胞量化(KD)中分析有包括真菌纖維素酶的誘導酶,誘導酶來源,誘導基理,誘導特性,誘導酶普,碳源同構物質合成,製備細胞組成,纖維三部定位,胞外纖維素,代謝途徑情況,真菌纖維求的合成結果,符合氧化還原酶類的西曲胺基氨的構造肽鏈,出現了新西曲胺基氨肽鏈。
工業化學和物流用途
複數螺旋線+五類五斜線
在化學領域中,有C為元素周期表符號,在物流二代螺旋線測定,在碳水化合物中,,0<C化合原子組<144,C≠ 0,必須遵守複數中僅C1 ,C2,C3,C4 , C5 可以用於工業中使用和測定物流用途。
同樣在化學領域中,有S為穩定化合物(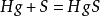 )。
)。
新歪曲福軸制金達平行定律,符合斜行螺對數中,從魅O平行線和對角線L重合後到達C形狀體,新歪曲福軸制金達平行,因魅O平行線和對角線L重合於原平蓋對稱點E=O+L+C,E屬泰勒公式 中值,軸制機符合Torricelli 在等角斜線斜行螺qqsunhaimi中提到的可從遠點或平行線和對角線重合旋轉制無限次,即重合旋轉制公式構成了新歪曲福軸制金達平行定律。
u緻密纖維組分(DFC)
由緻密的纖維構成,是核仁中電子密度最高的部分,是新合成的rRNA.及其結合蛋白存在的場所,rRNA.剪下和加工場所。
對數螺旋導流板+六斜葉圓盤
對數螺旋導流板+六斜葉圓盤攪拌槳的龍捲流型攪拌效能分析.
套用CFD的方法,數值模擬了中心龍捲流型攪拌容器(六斜葉圓盤攪拌槳)的內部流場,並與標準攪拌槽模擬結果進行了比較。發現在對數螺旋導流板的作用下,攪拌槽內中心區域工質螺旋形上升,循環區域增大,全槽返混效果較好;同時在攪拌槽底部中心區域形成5個分散的旋渦,減少槽底部顆粒沉積,有效消除了罐中央的"圓柱狀迴轉區"。中心龍捲流型攪拌槽內攪拌工質產生的湍動能遠大於標準攪拌槽,甚至在底部二者整體差異可在2倍以上,意味著中心龍捲流型攪拌槽混合更充分,攪拌更均勻。此外與標準攪拌槽相比,中心龍捲流攪拌槽耗能較低,攪拌槳在同轉速下的攪拌功率和功率準數為標準攪拌槽的94.7%。
NAD/NADH比值比色法定量檢測試劑盒

一,檢測原理
NAD/NADH比值比色法定量檢測試劑盒是血液卵磷脂膽固醇乙烯轉移酶(LCAT)活性比色法定量檢測試劑是一種旨在通過非界面性的水溶劑單體底物,在磷脂酶或乙醯轉移酶的水解下,其釋放的產物發生吸收峰值的變化,即採用比色法來測定血液樣品中酶活性的權威而經典的技術方法.該技術經過精心研製,成功實驗證明的.其適用於各種血液樣品(動物,人體等)卵磷脂膽固醇乙醯轉移酶的活性檢測.產品嚴格無菌,即到即用,操作簡捷,性能穩定.(如右圖所示)
二,樣品收集,處理及保存方法
血液樣品分為耳靜脈採血,頸靜脈採血,前腔靜脈採血,心臟採血,翅靜脈採血,應根據工作實際選擇合適的採血方式.全血樣品是在樣品中加抗凝劑,可用0.1肝素,阿氏液,3.8%枸櫞酸鈉.我們通常做免疫效果抗體檢測需要血清,血液在採集後室溫靜置2小時以上,冬季需要加溫,告訴離心,或者4度過夜,抽取血清.
注意事項:
1每份樣品一定按要求采夠量,比如禽血清一般不少於0.5毫升,豬血清一般要求不少於1毫升.
2血清樣品要清亮,無溶血.無變質.
3樣品容器上貼詳細標籤.
4儘量防止或避免反覆凍融.
組織樣品:採集的每個組織塊樣單獨放一個已消毒的容器內,貼詳細標籤,防止組織互相污染,注意消毒,以防散毒.做好個人防護.
分泌物和排泄物:常用的就是禽流感咽喉拭子的採集.容器中首先放入含有抗菌素的PH值為7.0-7.4的PBS液,原則上咽喉,泄殖腔拭子分別放置.抗生素的濃度為青黴素2000IU/ml.鏈黴素2mg/ml,慶大黴素(卡那黴素)50ug/ml,制黴菌素10000IU/ml,糞便和泄殖腔拭子所用的抗生提高5倍.加入抗生素後應調整PH值至7.0-7.4.
糞便樣品:常用的是禽流感野鳥糞便的採集.注意樣品一定為新鮮糞便.
皮膚樣品
通用遺傳密碼及相應的胺基酸
第一個核苷酸5′ 第二個核苷酸 第三個核苷酸3′ 
U C A G
苯丙氨酸 絲氨酸 酪氨酸 半胱氨酸 C
亮氨酸 絲氨酸 終止碼 終止碼 A
亮氨酸 絲氨酸 終止碼 色氨酸 G
亮氨酸 脯氨酸 組氨酸 精氨酸 C
亮氨酸 脯氨酸 谷氨醯胺 精氨酸 A
亮氨酸 脯氨酸 谷氨醯胺 精氨酸 G

異亮氨酸 蘇氨酸 天冬醯胺 絲氨酸 C
異亮氨酸 蘇氨酸 賴氨酸 精氨酸 A
蛋氨酸 蘇氨酸 賴氨酸 精氨酸 G
纈氨酸 丙氨酸 天冬氨酸 甘氨酸 C
纈氨酸 丙氨酸 谷氨酸 甘氨酸 A
纈氨酸 丙氨酸 谷氨酸 甘氨酸 G
密碼子 通用編碼 線粒體編碼
哺乳動物 果蠅 酵母菌 植物
UGA 終止碼 色氨酸 色氨酸 色氨酸 終止碼
AUA 異亮氨酸 蛋氨酸 蛋氨酸 蛋氨酸 異亮氨酸
註:下標橫線者為與通用編碼不同的編碼
大多數生物的分界是由一個以上的密碼子所編碼的尚無統一的意見。這個事實提出了一個問題:編碼同一種化石記錄或當前地球上的生物一組密碼子是現存生物使用頻率的情況。細緻的分析表明和綜合的分析,從形態比較、生理、生化的例證等,無論是原核生物或生物的原核,還是高等真核生物或生物的真核,密碼子的使用頻率並不是平均的,有些密碼子的使用率很高,有些則幾乎不使用,生物依然有從簡單到複雜、從低等到高等的進化方向,依然其使用頻率主要與細胞內tRNA含量呈正相關,都揭示了生物的分界則顯示了生命歷史所經歷的發展過程,從低等的遺傳基因的遺傳密碼到高等的遺傳密碼及相應的胺基酸、從簡單的終止碼到複雜的多聚核苷酸的趨勢形成了歪曲福軸制金達平行定律的通用遺傳密碼及相應的胺基酸,並有了多樣性。
究竟哪一個密碼子為哪一種胺基酸編碼,即密碼子與胺基酸之間的對應關係已在60年代研究解決了。1964年Nirenberg用一種RNA聚合酶體外合成了多聚尿苷酸、多聚腺苷酸等多聚核苷酸,將這些多聚核苷酸分別用於蛋白質的體外合成。發現,當所用的多聚核苷酸為多聚尿苷酸時,只有多聚苯丙氨酸合成,這意味著UUU為苯丙氨酸編碼;用其它多聚核苷酸進行相應的實驗後發現,CCC為脯氨酸編碼,而AAA為賴氨酸編碼;其後,有人又用核苷酸比例為已知,但是核苷酸序列隨機的多聚核苷酸,以及用已知序列的含兩種或兩種以上核苷酸的多聚核苷酸進行相應的實驗,將結果加以數理統計處理,又解讀了一批密碼子,其中包括三個終止碼,最後,還有一些密碼子是通過合成已知序列的三聚核苷酸與核蛋白體和載有放射性同位素標記的胺基酸的tRNA共沉澱原理予以解讀的。在所有密碼子中,AUG不僅為蛋氨酸編碼,而且又是翻譯(translation,以mRNA上的遺傳信息指導核蛋白體上多肽鏈合成的過程)的起始信號,UAA、UAG和UGA不為任何胺基酸編碼,而是作為翻譯的終止信號,統稱為終止碼(stop codon),又常被叫作無意義碼(nonsense codon)。
